Ingeniørbaserte modeller kan bedre forklare naturens mønster enn felles avstamning
Oversatt herfra.
 I tidligere artikler har jeg beskrevet hvordan ingeniørprinsipper bedre forklarer tilpasning enn evolusjonsteorien gjør (her, her, her, her). Nå skal jeg forklare hvordan ingeniørbaserte modeller også bedre forklarer mønsteret av likheter og forskjeller mellom arter i de høyere taksonomiske gruppene (f.eks. phyla, klasser og ordener) enn teorien om felles avstamning.
I tidligere artikler har jeg beskrevet hvordan ingeniørprinsipper bedre forklarer tilpasning enn evolusjonsteorien gjør (her, her, her, her). Nå skal jeg forklare hvordan ingeniørbaserte modeller også bedre forklarer mønsteret av likheter og forskjeller mellom arter i de høyere taksonomiske gruppene (f.eks. phyla, klasser og ordener) enn teorien om felles avstamning.
Dataforsker Winston Ewert demonstrerte at fordelingen av de samme genfamiliene i forskjellige arter passer langt bedre til det han omtaler som en avhengighets-grafmodell enn den vanlige avstamnings-modellen -Lenke. Hans sentrale tese er at likheter i livet representerer moduler som ble implementert i forskjellige arter for å oppnå lignende mål. Denne spådommen har blitt validert av flere forskningslinjer de siste tiårene.
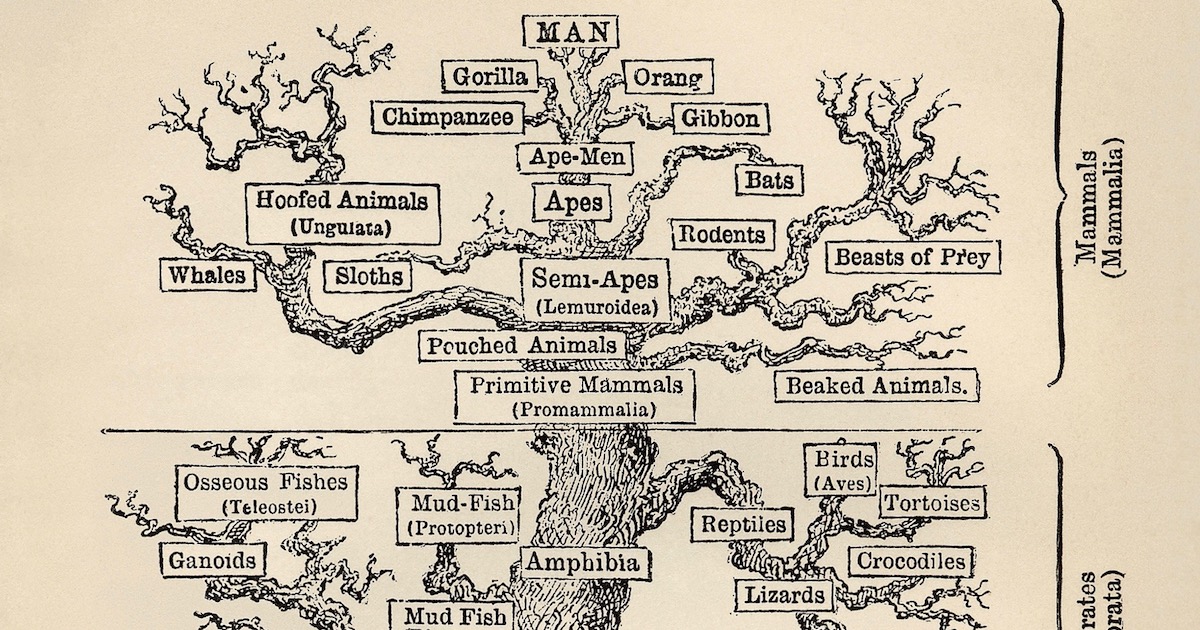
Kollapsen til livets tre
Jeg har tidligere beskrevet hvordan evolusjonister presenterer det påståtte livets tre (LT) som deres sterkeste bevis for felles aner -lenke, og jeg forklarte hvorfor LT kollapser (se, "BIO-kompleksitet presenterer en bedre modell enn felles aner for å forklare naturens mønster." -Lenke )
Som en synopse viser forskning på de Reskalerte Konsistens-Indeksene (RKI) for en rekke dyregrupper den sjokkerende inkonsistensen mellom prediksjonen om felles aner og det faktiske mønsteret av likheter i forskjellige grupper av arter. En RKI på 1 indikerer at alle  dataene passer med et evolusjonstre. En RKI på 0 indikerer at fordelingen av likheter ikke passer bedre til et tre enn helt tilfeldige data.
dataene passer med et evolusjonstre. En RKI på 0 indikerer at fordelingen av likheter ikke passer bedre til et tre enn helt tilfeldige data.
For å nevne noen få eksempler -lenke, ga data om egenskaper hos leddyr en RKI på 0,39. Den beregnede RKI for terapsider (påståtte forfedre til pattedyr) var 0,42. RKI for primater og deres påståtte nærmeste slektninger var 0,29. Og RKI for hvaler (f.eks. hvaler og delfiner) og deres påståtte nærmeste slektninger var 0,24. Evolusjonister siterer disse gruppene som å representere noen av de sterkeste bevisene for felles aner, men deres lave RKIs vitner om det motsatte.
Evolusjonister spådde historisk at RKI-verdier ville komme langt nærmere 1 enn 0, noe de tydeligvis ikke gjør. De lave verdiene viser at antakelsen om at likheter i egenskaper pålitelig antyder at felles avstamning er falsk. Problemet har blitt så akutt for mikroorganismer at mikrobiologene Vicky Merhej og Didier Raoult skildrer forsøk på å identifisere LT i de mest nedsettende termer (Merhej og Raoult, 2012):
-Ingen av de syv punktene som er angitt i introduksjonen til dette manuskriptet kan beholdes permanent, slik det ble etablert av Darwins teori, som på den tiden var en kamp mot kreasjonistene. Denne teorien kan ikke opprettholdes i sin helhet. Nylige fremskritt fra genomikk motbeviser ideene om gradualisme, eksklusiv vertikal arv, evolusjonsvalg av de sterkeste, en felles stamfar og TOL. Faktisk er det kanskje ikke to gener som har det samme evolusjonstreet.
Skuffende resultater
 Disse skuffende resultatene har tvunget evolusjonister til å foreslå flere mekanismer for å forklare de allestedsnærværende inkonsekvensene. Eksempler inkluderer Lateral GenOverføring (LGO), differensielt gentap, ufullstendig avstamnings-sortering og konvergent evolusjon. LGO refererer til gener som går fra en organisme til en annen. Denne prosessen kan teoretisk forklare hvordan de samme genene opptrer i ubeslektede arter, men sannsynligheten for utbredt LGO i komplekse organismer har blitt satt alvorlig spørsmålstegn ved -lenke. Ufullstendig avstamningssortering og gentap kan ikke forklare komplekse egenskaper som vises i fjernt beslektede organismer. Og påstanden om at komplekse tilpasninger kan utvikle seg uavhengig flere ganger (dvs. konvergent evolusjon) kollapser ved nøye undersøkelse på grunn av usannsynligheten av at de vises gjennom ikke-styrte prosesser selv én gang.
Disse skuffende resultatene har tvunget evolusjonister til å foreslå flere mekanismer for å forklare de allestedsnærværende inkonsekvensene. Eksempler inkluderer Lateral GenOverføring (LGO), differensielt gentap, ufullstendig avstamnings-sortering og konvergent evolusjon. LGO refererer til gener som går fra en organisme til en annen. Denne prosessen kan teoretisk forklare hvordan de samme genene opptrer i ubeslektede arter, men sannsynligheten for utbredt LGO i komplekse organismer har blitt satt alvorlig spørsmålstegn ved -lenke. Ufullstendig avstamningssortering og gentap kan ikke forklare komplekse egenskaper som vises i fjernt beslektede organismer. Og påstanden om at komplekse tilpasninger kan utvikle seg uavhengig flere ganger (dvs. konvergent evolusjon) kollapser ved nøye undersøkelse på grunn av usannsynligheten av at de vises gjennom ikke-styrte prosesser selv én gang.
For eksempel antas øyne med linser å ha utviklet seg uavhengig flere ganger, men alle evolusjonære scenarier møter uoverstigelige barrierer. De første mutasjonene i opprinnelsen til en linse ville allokere vev foran fotoreseptorene. Men udifferensiert vev ville hindre lysmottak, så de første mutasjonene ville raskt forsvinne. Linsen ville ikke bli gunstig før en kompleks utviklingsprosess koblet til et nytt genregulerende nettverk dukket opp -lenke. Likevel er den tilgjengelige tiden basert på fossilregistreringen utilstrekkelig til at selv den minste mengde nødvendig ny genetisk informasjon kan oppstå (her, her, her).
Ingeniørmoduler
 Derimot forklarer ingeniørbaserte modeller godt mønsteret av likheter gjennom livet. Designarkitekturer faller ofte inn i et hierarkisk mønster. Alle transportkjøretøyer har visse fellestrekk som tildelt plass for last og/eller passasjerer, et fremdriftssystem og styring. Biler har alle disse funksjonene pluss komponenter som hjul, bremser, kjølevæske, smøremidler og aksler. Toyota Camry-modeller har alle disse funksjonene pluss ytterligere spesialiserte komponenter. Likhetene i transportkjøretøyer vil sannsynligvis passe inn i et konstruert tre minst like godt som for forskjellige grupper av arter.
Derimot forklarer ingeniørbaserte modeller godt mønsteret av likheter gjennom livet. Designarkitekturer faller ofte inn i et hierarkisk mønster. Alle transportkjøretøyer har visse fellestrekk som tildelt plass for last og/eller passasjerer, et fremdriftssystem og styring. Biler har alle disse funksjonene pluss komponenter som hjul, bremser, kjølevæske, smøremidler og aksler. Toyota Camry-modeller har alle disse funksjonene pluss ytterligere spesialiserte komponenter. Likhetene i transportkjøretøyer vil sannsynligvis passe inn i et konstruert tre minst like godt som for forskjellige grupper av arter.
Mens mange funksjoner i menneskelige kreasjoner passer inn i et hierarkisk trelignende mønster, bryter mange det mønsteret. En politibil og et fly har begge toveis radioer mens toveis radioer er fraværende i de fleste andre biler. I tillegg er de samme kretsene implementert i en lang rekke kjøretøy for å møte lignende mål. Dette mønsteret gjenspeiler hvordan ingeniører ofte lager moduler de implementerer i ulike sammenhenger. Modulene må være utformet med den eksplisitte hensikten å operere i forskjellige produkter, og produktene som bruker modulene må være utformet for å integrere dem på riktig måte i deres operasjoner. Det samme mønsteret og begrensningene finnes i livet.
Mange av de samme egenskapene er implementert i forskjellige skapninger for å utføre lignende funksjoner. Ulike versjoner av øyne (f.eks. sammensatte og kameralignende) vises uavhengig flere ganger for å tillate optimalt syn i hver arts spesielle kontekst. Og svært like nevrale og utviklingsmoduler vises uavhengig i urelaterte arter. Nevrovitenskapsmenn Sanes og Zipursky identifiserte -lenke, i både fluers og menneskelige visuelle systemer bemerkelsesverdig like designmotiver i netthinnen, nevrale kretsløp i hjernen og genetiske kontrollmekanismer under utvikling, selv om de forskjellige øynene antas å ha utviklet seg uavhengig -lenke.
"Robust perfekt tilpasning"
På samme måte demonstrerte matematiske biologer Robyn Araujo og Lance Liotta -lenke; at alle biologiske nettverk som utfører "Robust Perfekte Tilpasninger" (RPT) kan dekomponeres i to distinkte klasser av moduler. Forskerne definerer RPT som "evnen til et system til å generere en utgang som går tilbake til et fast referansenivå (dets 'settpunkt') etter en vedvarende endring i input-stimulus, uten behov for justering av systemparametere." De samme modulene vises i systemer så forskjellige som signaloverføring, genregulering, protein-interaksjonsnettverk, sensoriske systemer og utviklingsregulering. De kombineres ofte hierarkisk for å utføre mer komplekse funksjoner.
 Av sentral viktighet opererer basismodulene innenfor strenge begrensninger:
Av sentral viktighet opererer basismodulene innenfor strenge begrensninger:
Komponentene deres må kobles sammen i henhold til spesifikke oppsett/maler (blueprints).
De kan ikke deles i undergrupper, så de består av et sett med komponenter som er irreduserbart komplekse.
Deres operasjoner må løse en lokal tilpasningsligning, så reaksjoner eller andre prosesser må operere innenfor stramme rammer.
Integreringen av modulene i større systemer må også oppfylle rigide matematiske kriterier.
Disse begrensningene innebærer at modulenes opprinnelse og implementering ikke kunne ha skjedd trinnvis gjennom en ikke-styrt prosess, siden deres konstruksjon og integrasjon krever koordinering, framsyn og målretning. Forfatterne av System Modeling in Cellular Biology (SMCB) -lenke, antar at evolusjon må være sann som en trosforpliktelse, men de kommenterer likevel:
.. konseptet "modulært design" er lånt fra menneskelig ingeniørkunst og har derfor en i hovedsak fremtidsrettet, målorientert natur. Komplekse motorer og nettverk er konstruert av moduler mens den endelige allmenne oppførselen til systemet holdes i bakhodet.
Mellom en ubåt og en fly
Naturmønsteret samsvarer også med ingeniørprinsipper i fordeling av artsmangfold. I menneskelig konstruksjon lages ofte mange versjoner av et bestemt design, for eksempel Toyota som produserer samme bilmodell med forskjellige sett med alternativer. Men få produkter ville falle utenfor de større kategoriene. Kjøretøy, som ville falle halvveis mellom en ubåt og et fly, har sjelden blitt designet.
På samme måte eksisterer det mange forskjellige arter som representerer forskjellige versjoner av samme tema. Det er identifisert flere ape- og menneskelignende skapninger som deler forskjellige likheter. Men likhetene passer ikke innenfor et konsekvent evolusjonstre. Og klart mellomliggende skapninger mellom primater og andre ordener av pattedyr har aldri blitt identifisert. Oppsummert, alle aspekter av mønsteret av likheter i naturen -motsier spådommene om felles avstamning, og passer til forventningene om ingeniørmodeller.
Oversettelse og bilder ved Asbjørn E. Lund